Seth Walsh

The man in the mirror is my only threat
Contributor
- Joined
- Jan 12, 2020
- Posts
- 9,504
- Reputation
- 18,742
Gene Expression Analysis
Fibrinogen Suppression (needed to create blood clots)
|
GHK also suppresses the production of the inflammatory cytokine interleukin-6 (IL-6) which is a main positive regulator of fibrinogen synthesis, through its interaction with fibrinogen genes. In cell culture systems, GHK suppresses IL-6 secretion in skin fibroblasts and IL-6 gene expression in SZ95 sebocytes.
In summary, the effects of GHK on the FGB gene plus its effects on IL-6 production imply a suppression of overall fibrinogen production.
Ubiquitin/Proteasome System
GHK stimulated gene expression in 41 UPS genes while suppressing only 1 UPS gene.
Up | Gene title | Percent change in gene expression |
1 | Ubiquitin specific peptidase 29, USP29 | 1056% |
2 | Ubiquitin protein ligase E3 component n-recognin 2, UBR2 | 455% |
3 | Gamma-aminobutyric acid (GABA) B receptor, 1 /// ubiquitin D, GABBR1 /// UBD | 310% |
4 | Ubiquitin specific peptidase 34, USP34 | 195% |
5 | Parkinson protein 2, E3 ubiquitin protein ligase (parkin), PARK2 | 169% |
6 | Ubiquitin-conjugating enzyme E2I (UBC9 homolog, yeast), UBE2I | 150% |
7 | Ubiquitin protein ligase E3 component n-recognin 4, UBR4 | 146% |
8 | Ubiquitin protein ligase E3B, UBE3B | 116% |
9 | Ubiquitin specific peptidase 2, USP2 | 104% |
10 | Ubiquitin-like modifier activating enzyme 6, UBA6 | 104% |
11 | Ubiquitination factor E4B (UFD2 homolog, yeast), UBE4B | 97% |
12 | Ubiquitin-conjugating enzyme E2M (UBC12 homolog, yeast), UBE2M | 92% |
13 | Ubiquitin-like modifier activating enzyme 7, UBA7 | 88% |
14 | HECT, C2 and WW domain containing E3 ubiquitin protein ligase 1, HECW1 | 81% |
15 | Proteasome (prosome, macropain) 26S subunit, ATPase, 3, PSMC3 | 81% |
16 | Ubiquitin-conjugating enzyme E2D 1 (UBC4/5 homolog, yeast), UBE2D1 | 79% |
17 | Proteasome (prosome, macropain) subunit, beta type, 2, PSMB2 | 79% |
18 | Ubiquitin protein ligase E3 component n-recognin 5, UBR5 | 77% |
19 | Ubiquitin specific peptidase 21, USP21 | 76% |
20 | OTU domain, ubiquitin aldehyde binding 2, OTUB2 | 76% |
21 | Proteasome (prosome, macropain) inhibitor subunit 1 (PI31), PSMF1 | 75% |
22 | Ubiquitin-conjugating enzyme E2H (UBC8 homolog, yeast), UBE2H | 73% |
23 | Ubiquitin-conjugating enzyme E2N (UBC13 homolog, yeast), UBE2N | 72% |
24 | Ubiquitin carboxyl-terminal hydrolase L5, UCHL5 | 71% |
25 | Proteasome (prosome, macropain) 26S subunit, non-ATPase, 13, PSMD13 | 70% |
26 | Ubiquitin associated protein 1, UBAP1 | 70% |
27 | Ubiquitin-conjugating enzyme E2B (RAD6 homolog), UBE2B | 69% |
28 | TMEM189-UBE2V1 readthrough /// ubiquitin-conjugating enzyme E2 variant 1, TMEM189-UBE2V1 /// UBE2V1 | 67% |
29 | Proteasome (prosome, macropain) 26S subunit, non-ATPase, 1, PSMD1 | 64% |
30 | Proteasome (prosome, macropain) 26S subunit, non-ATPase, 3, PSMD3 | 64% |
31 | Ariadne homolog, ubiquitin-conjugating enzyme E2 binding protein, 1 (drosophila), ARIH1 | 61% |
32 | BRCA1 associated protein-1 (ubiquitin carboxy-terminal hydrolase), BAP1 | 60% |
33 | Ubiquitin interaction motif containing 1, UIMC1 | 60% |
34 | Ubiquitin associated protein 2-like, UBAP2L | 57% |
35 | Ubiquitin protein ligase E3 component n-recognin 7 (putative), UBR7 | 56% |
36 | Ubiquitin-conjugating enzyme E2G 1 (UBC7 homolog, yeast), UBE2G1 | 54% |
37 | Itchy E3 ubiquitin protein ligase homolog (mouse), ITCH | 54% |
38 | Ubiquitin-conjugating enzyme E2D 4 (putative), UBE2D4 | 51% |
39 | Proteasome (prosome, macropain) 26S subunit, non-ATPase, 10, PSMD10 | 50% |
40 | WW domain containing E3 ubiquitin protein ligase 1, WWP1 | 50% |
41 | Ubiquitin-like 3, UBL3 | 50% |
Down | Gene title | Percent change in gene expression |
1 | Ubiquitin associated and SH3 domain containing A, UBASH3A | −89% |
DNA Repair Genes
GHK was primarily stimulatory for DNA repair genes (47 UP, 5 DOWN).
|
The most affected DNA repair genes.
|
Antioxidant Genes
Among the 13,424 available genes in the Broad Institute data, we were able to identify 14 antioxidant genes in which GHK stimulates as well as two prooxidant genes that GHK suppresses. GHK increases the expression of the oxidative/inflammatory gene NF-κB2 103% but also increases the expression of two inhibitors of NF-κB, TLE1 by 762% and IL18BP by 295%, thus possibly inhibiting the activity of the NF-κB protein.
Up | Genes | Percent change in gene expression | Comments |
1 | TLE1 | 762% | Inhibits the oxidative/inflammatory gene NF-κB. |
2 | SPRR2C | 721% | This proline-rich, antioxidant protein protects outer skin cells from oxidative damage from ROS. When the ROS level is low, the protein remains in the outer cell membrane but when the ROS level is high, the protein clusters around the cell’s DNA to protect it. |
3 | ITGB4 | 609% | Upregulation of ITGB4 promotes wound repair ability and antioxidative ability. |
4 | APOM | 403% | Binds oxidized phospholipids and increases the antioxidant effect of HDL. |
5 | PON3 | 319% | Absence of PON3 (paraoxonase 3) in mice resulted in increased rates of early fetal and neonatal death. Knockdown of PON3 in human cells reduced cell proliferation and total antioxidant capacity. |
6 | IL18BP | 295% | The protein encoded by this gene is an inhibitor of the proinflammatory cytokine IL18. IL18BP abolished IL18 induction of interferon-gamma (IFNgamma), IL8, and activation of NF-κB in vitro. Blocks neutrophil oxidase activity. |
7 | HEPH | 217% | Inhibits the conversion of Fe(2+) to Fe(3+). HEPH increases iron efflux, lowers cellular iron levels, suppresses reactive oxygen species production, and restores mitochondrial transmembrane potential. |
8 | FABP1 | 186% | Reduces intracellular ROS level. Plays a significant role in reduction of oxidative stress. |
9 | PON1 | 149% | PON1 (paraoxonase 1) is a potent antioxidant and a major anti-atherosclerotic component of high-density lipoprotein. |
10 | MT3 | 142% | Metallothioneins (MTs) display in vitro oxyradical scavenging capacity, suggesting that they may specifically neutralize hydroxyl radicals. Metallothioneins and metallothionein-like proteins isolated from mouse brain act as neuroprotective agents by scavenging superoxide radicals. |
11 | PTGS2 | 120% | Produces cyclooxygenase-II (COX-II) which has antioxidant activities. |
12 | NF-κB2 | 103% | NF-κB, an oxidative/inflammatory protein, is involved in cellular responses to stimuli such as stress, cytokines, free radicals, ultraviolet irradiation, oxidized LDL, and bacterial or viral antigens. |
13 | NFE2L2 | 56% | Nuclear respiratory factor 2 helps activate antioxidant responsive element regulated genes which contribute to the regulation of the cellular antioxidant defense systems. |
14 | PTGS1 | 50% | Produces cyclooxygenase-I (COX-I) which has antioxidant activity. |
Down | Genes | Percent change in gene expression | Comments |
1 | IL17A | −1018% | This strongly suppressed cytokine can stimulate the expression of IL6 and cyclooxygenase-2 (PTGS2/COX-2), as well as enhancing the production of nitric oxide (NO). High levels of this cytokine are associated with several chronic inflammatory diseases including rheumatoid arthritis, psoriasis, and multiple sclerosis (NCBI GENE entry). |
2 | TNF | −115 | GHK suppresses this prooxidant TNF gene which inhibits the antioxidant IL18. |
2.6. Insulin and Insulin-Like System
GHK stimulates 3 genes in this system and suppresses 6 genes.
Up | Gene title | Percent change in gene expression |
1 | Insulin-like 6, INSL6 | 188% |
2 | Insulin-like growth factor 2 mRNA binding protein 3, IGF2BP3 | 136% |
3 | Insulin-like growth factor binding protein 3, IGFBP3 | 82% |
Down | Gene title | Percent change in gene expression |
1 | Insulin-like growth factor 1 (somatomedin C), IGF1 | −522% |
2 | Insulin receptor-related receptor, INSRR | −437% |
3 | Insulin, INS | −289% |
4 | Insulin-like 3 (Leydig cell), INSL3 | −188% |
5 | Insulin-like growth factor binding protein 7, IGFBP7 | −110% |
6 | Insulin-like 5, INSL5 | −101% |
Summary:
Fibrinogen
Fibrinogen, the protein which is used to make blood clots, is also a strong predictor of mortality in cardiovascular patients. After vascular incidents, such as myocardial infarction, fibrinogen concentrations increase sharply. The free, unclotted fibrinogen protein increases the “stickiness” of red blood cells which stack together forming rouleaux. This increases the time of the “solid” blood state which decreases blood flow through the microcirculation where blood flows like a thixotropic fluid, switching between a solid phase and a liquid phase, somewhat like toothpaste. As a solid, it stops oxygen and nutrient flow to the tissues. This, in itself, can cause tissue damage.
The gene data on GHK’s suppression of FGB (the fibrinogen beta chain) combined with its actions on lowering IL-6 secretion on fibroblasts and sebocytes appears to be sufficient to explain its lowering effect on fibrinogen.
Ubiquitin Proteasome System
The ubiquitin proteasome system (UPS) functions in the removal of damaged or misfolded proteins. Aging is a natural process that is characterized by a progressive accumulation of unfolded, misfolded, or aggregated proteins. In particular, the proteasome is responsible for the removal of normal as well as damaged or misfolded proteins. Recent work has demonstrated that proteasome activation by either genetic means or use of compounds retards aging.
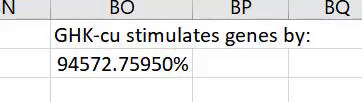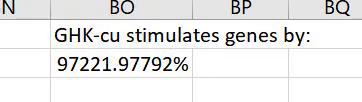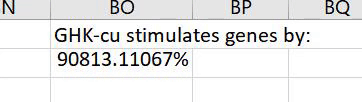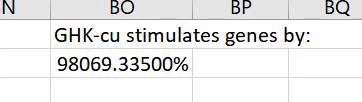
In our screening of UPS genes with a percent change of at least ±50%, GHK increased gene expression in 41 UPS genes while suppressing 1 UPS gene. Thus, it should have a positive effect on this system.
DNA Repair
It is estimated that normal metabolic activities and environmental factors such as UV light and radiation can cause DNA damage, resulting in somewhere between 1000 and as many as 1 million individual molecular lesions per cell per day. Lack of sufficient DNA repair is considered a cause of cell senescence, programmed cell death, and unregulated cell division, which can lead to the formation of a tumor that is cancerous.
GHK was stimulatory for DNA repair genes (47 stimulated, 5 suppressed) suggesting an increased DNA repair activity.
Antioxidant Defense
Free radicals and toxic end products of lipid peroxidation are linked to atherosclerosis, cancer, cataracts, diabetes, nephropathy, Alzheimer’s disease, and other severe pathological conditions of aging. Reactive oxygen species (ROS) and reactive carbonyl species (RCS) are produced in cells in small quantities under physiological conditions and play an important role in cell signaling and immune defense. A robust antioxidant network maintains balance between free radical production and scavenging, ensuring that the overall damage from free radicals is low. However, in the course of aging and in pathological conditions such as inflammation, the balance may shift toward free radical accumulation that can lead to oxidative stress and eventually to cell death.
GHK increases gene expression of 14 antioxidant genes and suppresses the expression of 2 prooxidant genes. It increases the expression of the oxidative/inflammatory gene NF-κB2 103% but also increases the expression of two inhibitors of NF-κB, TLE1 by 762% and IL18BP by 295%; thus, it possibly inhibits the activity of the NF-κB protein.
GHK also possesses antioxidant activities in cell culture and in vivo.
In dermal wound healing in rats, GHK, attached to biotin to bind it to collagen pads covering wounds, produced a higher production of protein antioxidants in the wound tissue. Superoxide dismutase was increased 80% while catalase was increased 56%. GHK reduced gastric mucosal damage by 75% against lipid peroxidation by oxygen-derived free radicals induced by acute intragastric administration of ethanol.
Interleukin 1 beta can induce serious oxidative damage to cultured cells. GHK markedly reduced oxidative damage by interleukin 1-beta to cultured insulin secreting pancreatic cells.
In another study, GHK entirely blocked the extent of in vitro Cu(2+)-dependent oxidation of low density lipoproteins (LDL). Treatment of LDL with 5 microM Cu(2+) for 18 hours in phosphate buffered saline (PBS) resulted in extensive oxidation as determined by the content of thiobarbituric acid reactive substances. Oxidation was entirely blocked by GHK. In comparison, copper, zinc-superoxide dismutase provided only 20% protection.
Acrolein, a well-known carbonyl toxin, is produced by lipid peroxidation of polyunsaturated fatty acids. GHK directly blocks the formation of 4-hydroxynonenal and acrolein toxins created by carbonyl radicals that cause fatty acid decomposition. GHK also blocks lethal ultraviolet radiation damage to cultured skin keratinocytes by binding and inactivating reactive carbonyl species such as 4-hydroxynoneal, acrolein, malondialdehye, and glyoxal.
Iron has a direct role in the initiation of lipid peroxidation. An Fe(2+)/Fe(3+) complex can serve as an initiator of lipid oxidation. The major storage site for iron in serum and tissue is ferritin and the superoxide anion can promote the mobilization of iron from ferritin which can catalyze lipid peroxidation. GHK : Cu(2+) produced an 87% inhibition of iron release from ferritin by apparently blocking iron’s exit channels from the protein.
Insulin and Insulin-Like Pathways
The insulin/IGF-1-like receptor pathway is a contributor to the biological aging process in many organisms. The gene expression data suggests that GHK suppresses this system as 6 of 9 of the affected insulin/IGF-1 genes are suppressed.
Insulin/IGF-1-like signaling is conserved from worms to humans. In vitro experiments show that mutations that reduce insulin/IGF-1 signaling have been shown to decelerate the degenerative aging process and extend lifespan in many organisms, including mice and possibly humans. Reduced IGF-1 signaling is also thought to contribute to the “antiaging” effects of calorie restriction.
Last edited: